
Research & Writings
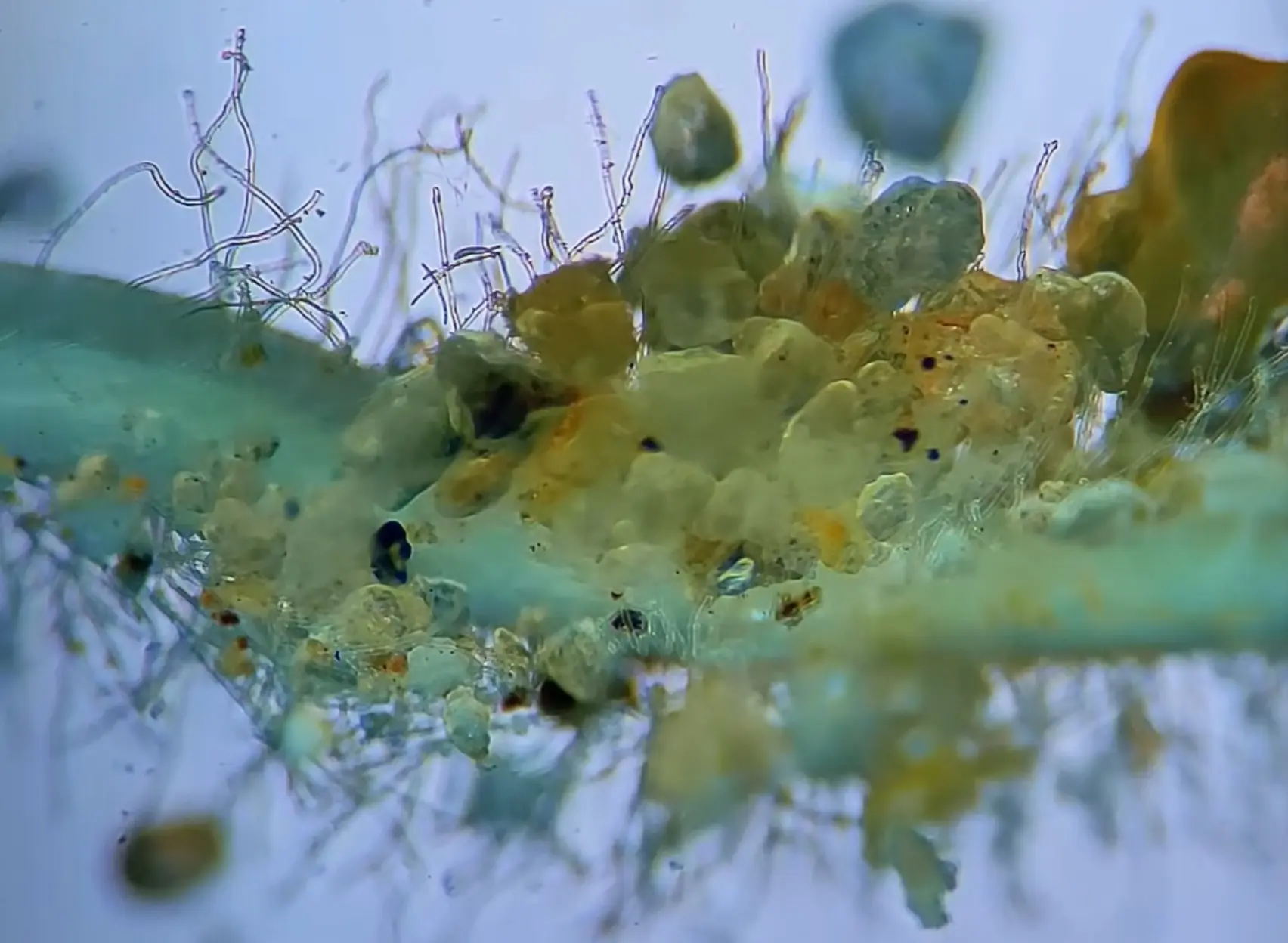
The Aquarium "Fungi"
Read Article →
The Tropical Fish Trap: Why "Tropical" is Not "Hot"
Read Article →
How Ecological Theory Fixes Aquarium Reality
Read Article →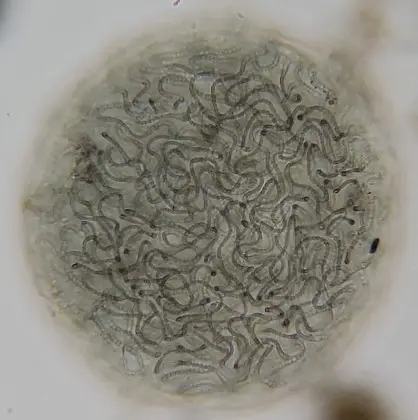
We’re Cycling Aquariums for Failure
Read Article →
SUBEX-2025: Substrate Comparison Experiment
View Findings →
Future Article about Anaerobic Pockets

Future Article about My 5 Years Old Nanocube
